@Arvind Arya - I live down in morgan hill but work up near great america in santa clara. DF will certainly lower the swimming populations - it takes ~300 throphants per gram of fish to be lethal - so the time to deploy DF in-tank would have been right when you first saw you had an issue. DF in the quarantine could still be of help.
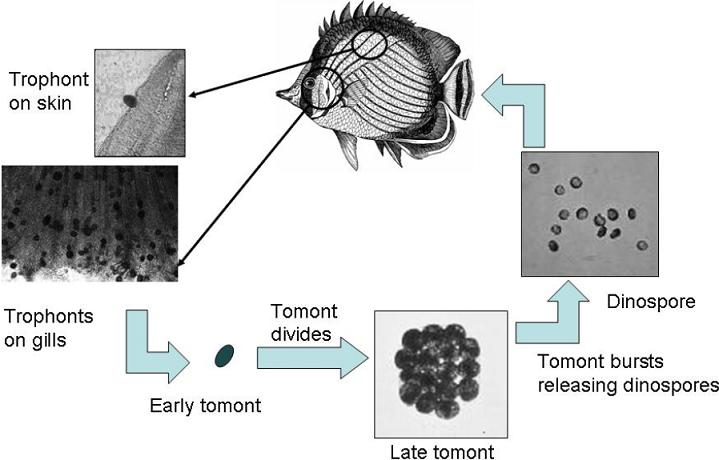
The infectivity of throphants is highly variable. Not all trophants find a fish, some that do can't attach and some of those do not successfully drop and adhere to produce tomonts.
Taking HF's numbers as a reference - even if you don't take out all free-swimmers and droppers - if you can reduce the infectivity rate below 200:1 - the parasitic cycle will break.
Working in your favor is the natural immunity building in the fish coupled with the senescence of the cell line. Keep the attachment rate below the lethal loading - and the parasite more or less self eradicates. It's simple math. DF also reduces bacterial load and helps prevent secondary infection - which is also a plus.
We tend to naively interpret lab results and try to apply them to our systems, sometimes without considering the fact that lab tanks are in many ways different from the tanks we run at home. There are many aspects of the life-cycle if ich and velvet that have not been explored and are not well understood but which are bit-by bit being figured out.
Senescence of cilliate lines was observed and described over a hundred years ago - and it is reliably observed in virtually every study of marine ich. In the lab - it can be a significant challenge to maintain a viable cell line - so much so that special techniques have had to be developed just to complete a study will a single strain before it dies off on its' own from old age even though it is "fed" a diet of fish which have never had any exposure and have acquired no immunity. Turns out there is a limit to how many times a cell can reproduce by budding as it typically takes place in the tomont under laboratory conditions. How is this observed senescence avoided in the wild - or as some here suggest - in our reef systems? No one knows for sure.
The are researchers looking for the genetic loci of ich's ability to enter into deep hibernation states. Of course they are looking based on its impact on fisheries and the observed seasonality of outbreaks. Is it entering or exiting these hibernation states, which revitalizes the cell line and restores infectivity? Sexual and/or autogamous reproduction which clears DNA damage and restores shortened telomeres - is that the missing piece of the life-cycle which keeps ich and velvet disappearing through senescent eradication? Looks possible. Looks probable as a matter of fact.
So what are the trigger(s) for deep hibernation? Are they the same for autogamous or sexual reproduction? What are the environmental conditions in a "lab" tank which precludes these states. We know temperature can have an effect - but if this were the only agent - then senescence should be the rule in our tanks as well. It turns out that there is (at least) one other environmental trigger. Hypoxic conditions with O2 levels < 23% have also been seen to be inductive of deep hibernation states. These regions do exist in our tanks - notably areas of the substrate. Tomonts of ich can be revived from hypoxic induced hibernation six, seven or eight months later after exposure to an oxygen rich environment. Thermoclines and/or hypoxia? Thermoclines in our systems - no.
Ich or velvet hibernating in sand beds or other regions of reduced 02 saturation - very possible.








") :
: 











